Wide world of venom - Scorpions
Venom is an adaptation that can be found in almost every major animal group, including fish, invertebrates, mammals, amphibians and of course reptiles. In a parallel blog series, Tim is contemplating a few of the fundamental questions and controversies that surround venom, such as which animals are venomous. If you have been following his posts, you will know by now that the answer is not as clear-cut as you might think (and if you haven’t been following them, I highly recommend you start!). However, for now we will leave the philosophising in Tim’s capable hands as we begin a series in which we showcase some of the diversity of venomous creatures (including a few of the contentious ones), their venom, and their venom delivery systems.

The crown-of-thorns starfish (Acanthaster planci) is venomous echinoderm. Image Derek Keats CC2.
The wide world of venom
The word “venom” tends to invoke feelings associated with fear and life-threatening situations in many people. However, as we are delving deeper into the world of venom, hopefully you are beginning to realise that this needn’t be the case. In fact, most venomous animals actually pose no real threat to us whatsoever. Venom has independently evolved across numerous lineages; it has even independently evolved within many lineages, such as at least 12 times within the spiny-rayed fishes (Acanthomorpha) alone.
You may remember from previous blogs that venom is broadly defined as “a toxic secretion that is produced by specialised glands or cells of one animal and delivered (via a specialised delivery mechanism) to another animal, through the infliction of a wound, which disrupts the normal physiological processes of the receiving animal and benefits the producing animal.” As we have bombarded you with a lot of information of late, let’s begin our journey with a quick recap of a few of these defining points.
A functional trait
The role of venom varies depending on the species, but it is usually employed in either a predatory or defensive context. This makes it a functional trait; a kind of evolutionary interface between organism and ecosystem, the use of which provides a direct contribution to the fitness of the venomous animal. In this way, venom may have played a key role in the explosive diversification of many of these venomous animal groups. That is, the emergence of venom probably gave these species a higher likelihood of success in adapting to and persisting in new and existing ecosystems, increasing their likelihood of survival and reproductive success. This is essentially the basis for speciation (the evolution of new species) and diversification (the rate at which new species evolve), which has resulted in a stunning diversity of venomous taxa that possess venom toxins and associated apparatus in an array of forms and functions.
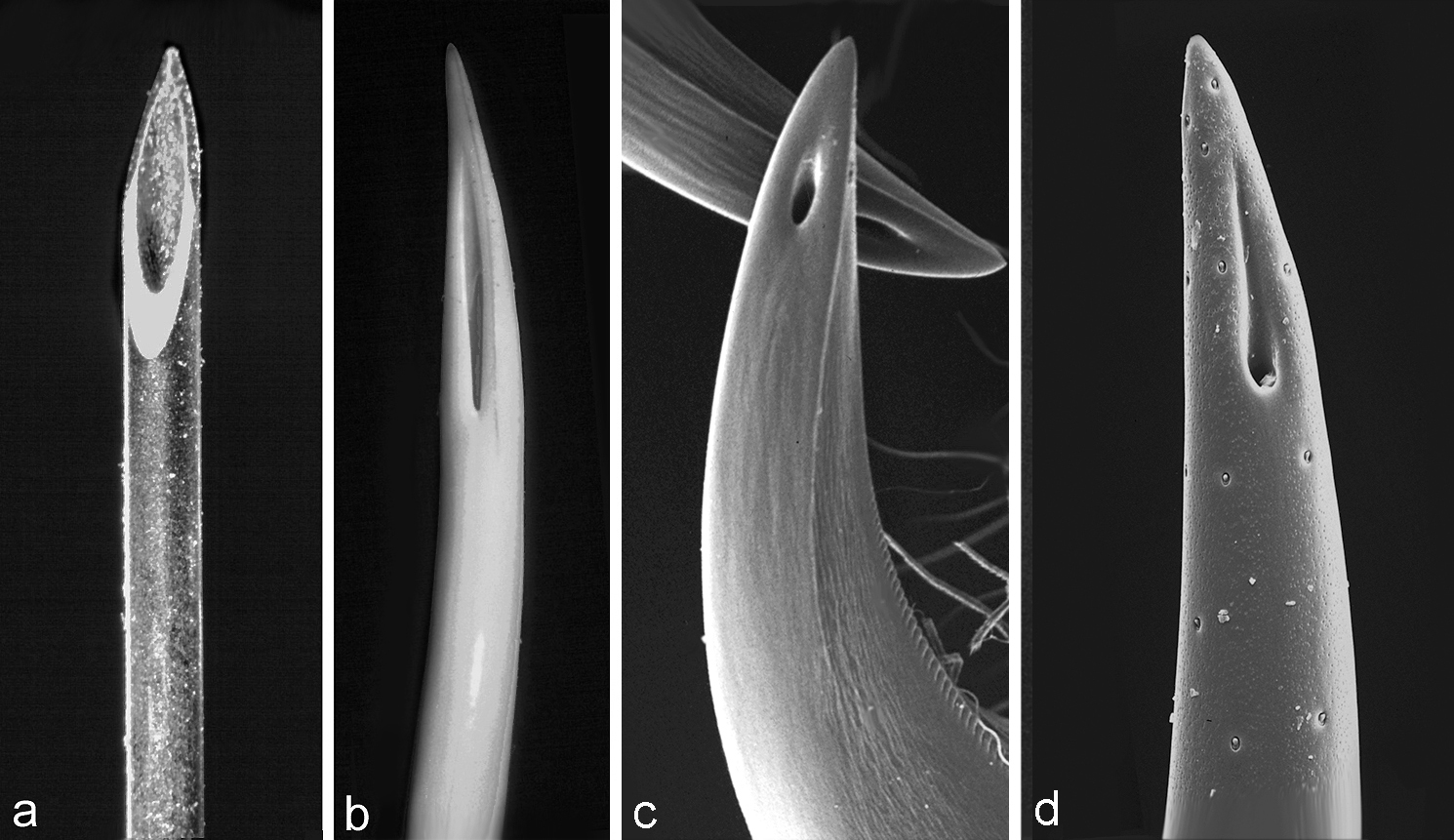
Some of the diversity and convergence in venom apparatus morphology is evident in this stunning image that compares: a) the tip of a hypodermic syringe; b) the tooth of a viper (Bitis gabonica); c) the cheliceral fang of a jumping spider (Portia sp.); d) lateral view of the stinger of a scorpion (Euscorpius italicus), in which only one of its two venom duct openings can be seen. People "invented" the hypodermic needle around 200 years ago, while the fang of the viper has been around for approximately 200 million years, that of the spider for around 300 million years, and the scorpion around 400 million years. It would seem we were a little late to the game! Image from Foelix 2011, Biology of Spiders.
It is not yet understood precisely how, genetically, venoms "came to be", or how toxins were (and continue to be) “recruited” for use as a biochemical weapon. However, DNA replication is an imperfect process, and mutations provide the perfect platform for the emergence of new, beneficial traits. Snake venom evolution represents perhaps the most extensively studied of any venomous group. Research indicates that most snake venom toxins are essentially derived from normal body proteins—digestive enzymes, for example—which, through some kind of genetic hiccup, were duplicated, expressed, and secreted by cells in oral glands (though not necessarily in that order), and were subsequently exapted (repurposed) and weaponised: delivered from one organism into the body of another, in which they exert a toxic effect (for a more in-depth discussion on these topics, I again refer you to Tim’s blog series!). Over evolutionary time, selection favoured mutated forms of the molecules that induced these toxic effects with more strength, speed, and/or efficiency. In parallel, specialised systems that facilitated the delivery of these secretions with greater efficiency developed, such as venom glands coupled with hollow or grooved fangs.

Many species of catfish have defensive venomous spines, but that doesn't always do the trick. Image Ellen & Tony CC2.
Research points toward a similar mechanism of recruitment in some other lineages. For example, investigations are underway to demonstrate the homology (shared ancestry) between the proteins secreted by the skin’s regulatory mucosal glands and those of the venom glands adjoining the spines in some species of catfish. The presence of such an integrated “venom system” (toxic secretion produced by specialised glands or cells, coupled with a specialised delivery mechanism) is considered by many scientists to be a key aspect of differentiating between animals that incidentally introduce their secretions into the body of another animal from those that intentionally do so. In conjunction, the adaptive value of the animal’s secretion (the measure of its usefulness once delivered) is also key. If it doesn’t benefit the producing animal once delivered, it isn’t really venom. For example, we humans have proteins in our oral secretions (saliva) that can also be found in venoms. We also have specialised glands producing these secretions. However, we do not have teeth specialised for injecting said secretions, nor do they offer us any adaptive value once out of our bodies. Therefore, we are not venomous. It is the way in which the toxins enter the victim’s body and how their bioactivity then serves a specific functional role that is crucial. This is also why many relatively harmless invertebrates, such as leeches and ticks, are widely considered to be venomous. Their specialised toxic secretions have a specific role in facilitating a certain behaviour of the venomous animal once they have been delivered into their target’s tissue, such as the anticoagulant toxins that facilitate the leech’s feeding.
Over the coming weeks, we are going to explore the diversity of the toxic secretions and associated apparatus of such animals in more detail. We will kick off the series by looking at the venom apparatus of the scorpions.
 A poised scorpion may invoke fear in many people, but this needn't necessarily be the case - most scorpion species pose no real threat to humans. But scorpions, like all other venomous and non-venomous creatures, should be treated with respect - this is the surest way to avoid any negative encounters. Image Sid Mosdell CC2.
A poised scorpion may invoke fear in many people, but this needn't necessarily be the case - most scorpion species pose no real threat to humans. But scorpions, like all other venomous and non-venomous creatures, should be treated with respect - this is the surest way to avoid any negative encounters. Image Sid Mosdell CC2.
Scorpions
The order Scorpiones is an ancient lineage of arachnid and its members are found in every continent except Antarctica. While a scorpion poised in its threat display with arching metasoma (tail) is an iconic symbol of venom, fewer than 30 of the 1700-ish species of scorpion are considered potentially lethal to humans.
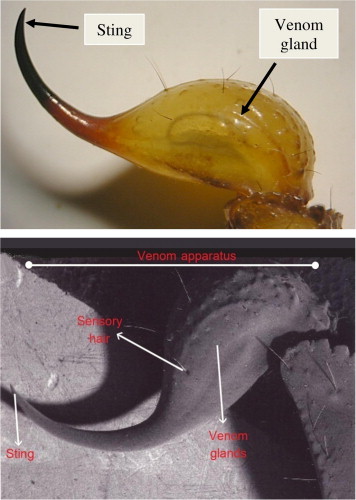
The venom apparatus of a scorpion forms the last segment of its metasoma (tail), comprising telson (bulbous region housing the venom gland) and aculei (stinger barb). Soliman, B. A., Shoukry, N. M., Mohallal, M. E., & Fetaih, H. A. (2013). Fine structure of the stinger, histology and histochemistry of the venom gland in the scorpion Androctonus amoreuxi (Buthidae). The Journal of Basic & Applied Zoology, 66(2), 41-46. CC3.
The venom system of a scorpion is comprised of paired venom glands housed within the bulbous telson (stinger), which is tipped by the aculeus (stinger barb). Their venom ducts are kept separate by a thin cuticle and porous layer, with two distinct openings near the tip of the aculeus. The telson is covered in sensory hairs, which may aid the scorpion in locating a softer area to sting its invertebrate prey. In contrast, the aculeus of the scorpion is smooth - likely to facilitate entry into predator or prey tissue - and is hardened via the accumulation of such heavy metals as zinc, iron, and manganese.
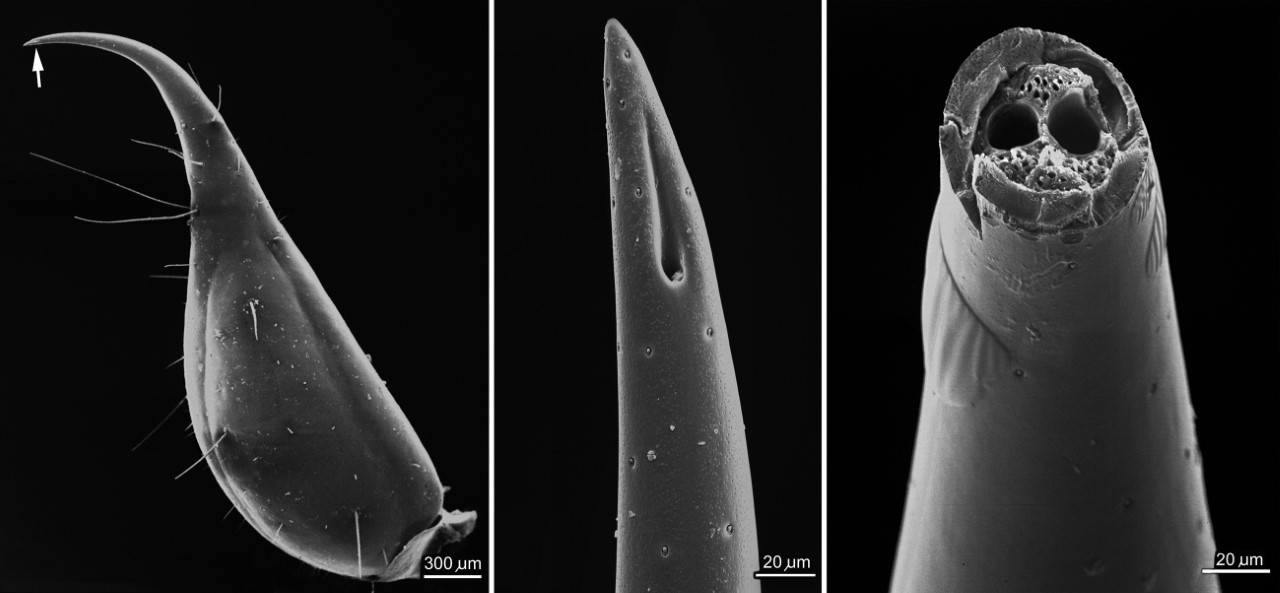
The telson, with arrow indicating site of venom duct opening (left); the aculeus tip showing venom duct opening (middle); and the interior of the aculeus showing paired venom ducts (right) of a Euscorpius. Image: RF Foelix.
Scorpion venoms are predominantly composed of neurotoxic peptides, meaning they disrupt nerve signalling - typically via ion-channel blockage. However, their venoms contain different components that bind with both vertebrate and invertebrate receptors, reflecting specialisation of toxin composition for use in both defence and predation. Though this behaviour can vary widely between species, it could be said that scorpions are some of few venomous creatures whose venom is specialised to act in both contexts. In fact, several members of the genus Parabuthus even spray their venom defensively, likely as a means of temporarily blinding their assailant. Scorpions also alter the composition of their venom to increase the concentration of toxins more suited to use in defence in as few as 6 weeks, following increased exposure to threat. Additionally, members of this same genus can eject a small quantity of “pre-venom” – a secretion with greater pain-inducing potential – before deploying a larger quantity of its “typical” venom.
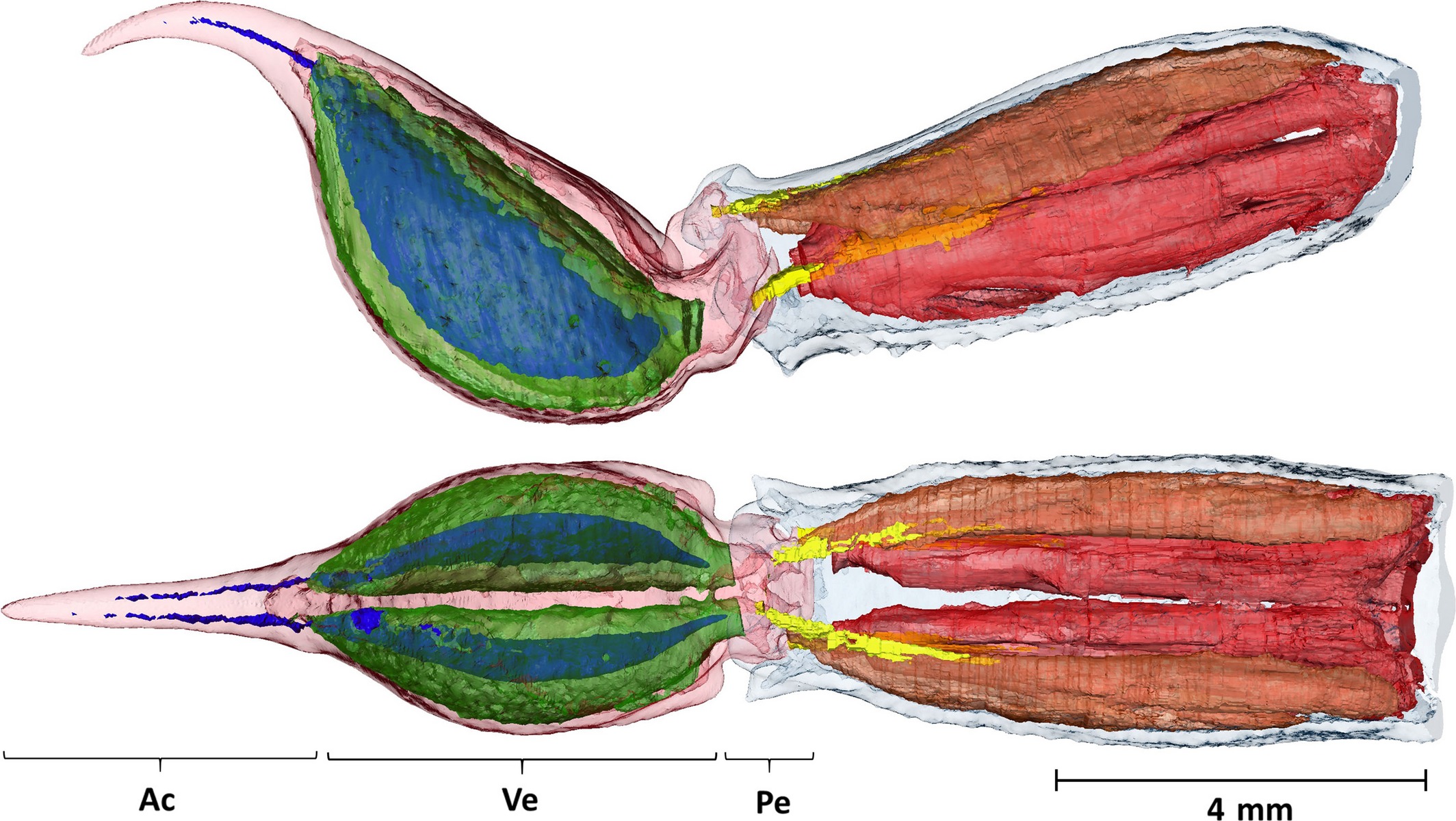
Internal anatomy of the telson and the fifth metasomal segment (Neochactas delicatus). Top: lateral view; bottom: dorsal view. Cuticula of the fifth metasomal segment rendered in transparent blue, cuticula of the telson rendered in transparent pink. The telson can be subdivided into the aculeus (Ac), vesicle (Ve) and peduncle (Pe). Telson shows the venom gland (transparent green) and venom canal lumen (blue) inside the vesicle. The discontinuous lumen of the venom ducts may be due to coagulated venom existing inside the ducts. The thin layer of musculature around the venom glands is not shown. The apodemes are in yellow and can be seen continuing inside the flexor (transparent brown) and extensor (transparent red) muscles. Note the larger volume of the extensor muscles, which allow a forceful extension of the telson in the direction of the aculeus tip. Scale bar: 4 mm. (van der Meijden, A., & Kleinteich, T. (2017). A biomechanical view on stinger diversity in scorpions. Journal of anatomy, 230(4), 497-509.A biomechanical view on stinger diversity in scorpions (2016) 30:4, 497-509. https://doi.org/10.1111/joa.12582. Reproduced by permission of Oxford University Press on behalf of The Society for Integrative and Comparative Biology.
Interestingly, many species of scorpion also demonstrate extreme variation in venom toxicity between males and females, the balance of which differs between species. However, given that some scorpions have been observed to alter their venom over a short timeframe, it is not known whether these are true sexual variations or reflections of venom plasticity. Male scorpions are also known to extensively sting their partners during courtship and mating. Whether envenoming occurs in parallel is not known, but the females apparently experience little ill-effect from stinging activity, and it has been suggested that differences in toxicity may be linked to this courtship stinging.

Though the size of scorpions' chelae (pincers) is linked to venom toxicity, this is not a reliable indicator. Image left: jbdodane CC2; right: Sarah Martin CC2.
The telson and chelae (pincers) of scorpions can vary substantially in size between genera. There is a crude inverse link between chelae size and venom toxicity; however, this is not a reliable indicator of toxicity, and chelae morphology is more indicative of differences in defensive and predatory stinging and grappling behaviour as opposed to venom. For example, some species are known to sting prey explosively, while others grapple and immobilise prey using their chelae and sting in a slow and deliberate manner once their prey has been firmly secured. Links have also been found to suggest that species with greater venom toxicity have longer and more curved aculei (stinger barbs). While the reason for this is unclear, it too is more likely related to stinging behaviour than venom.
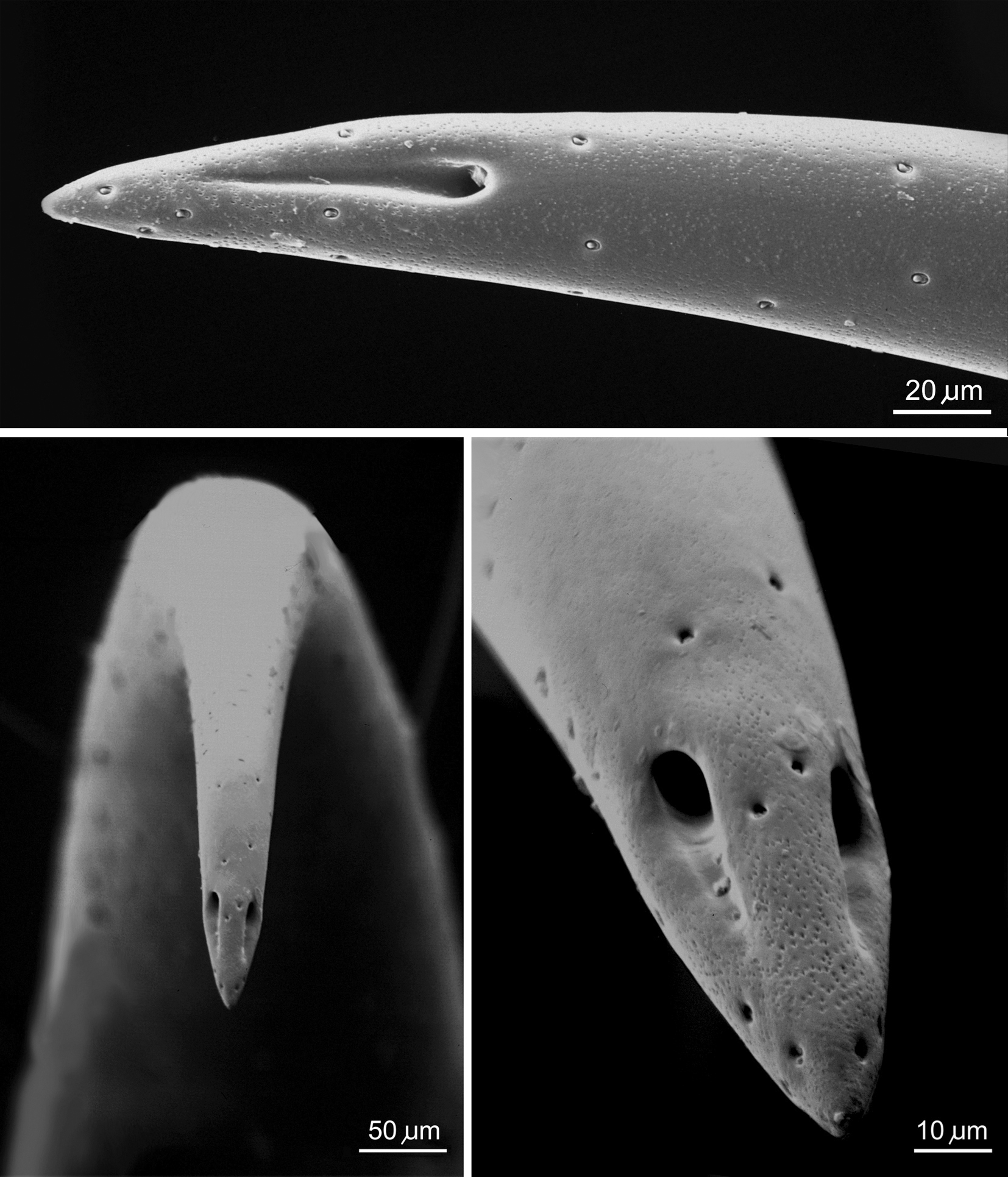
The curved aculeus with paired venom duct openings of a scorpion. Image: RF Foelix.
Thanks for joining us for part 1 of our series celebrating the diversity of venomous creatures. Next week, we will be looking at the venom systems of the spiny-rayed fishes (Acanthomorpha).