The tale of Toxicofera, part 3
Last week we discussed exaptation – the idea that certain features of organisms that are designed for a particular function may have side effects that make them suited to repurposing for another function. The simple example we used was feathers, which initially functioned as insulation, but were later repurposed for flight. The same kind of process is important for the evolution of all kinds of functional traits. Remember: you can’t get something from nothing; the future is built from the past. So, it stands to reason that this kind of repurposing (exaptation) is also important for the evolution of the functional trait that this blog series is all about – venom.
Being venomous is all about getting enough of a toxic substance you produce into the body of another organism, so that you can eat that organism or prevent it from eating you. So, there are two clear components to the venomous condition – a toxic secretion and a delivery mechanism. Exaptation is centrally involved in the origins of both of these components. Last week we talked a little bit about functions of oral secretions that make them good “proto-venoms” – antimicrobial activity, as well as pre-digestive activities (which we only mentioned in passing). Both of these involve breaking down molecules (including those which comprise cell walls). Since organisms are made of molecules, a good place to start if you want to subdue or deter one is breaking some of its molecules apart. In a future blog post, we’re going to have a much deeper look at the role of exaptation (again, think repurposing or “co-option”) in turning a protein with a particular function (either in saliva or somewhere else in the body) into a toxin. This week, however, we’re going to focus on the origins of reptilian venom delivery systems.
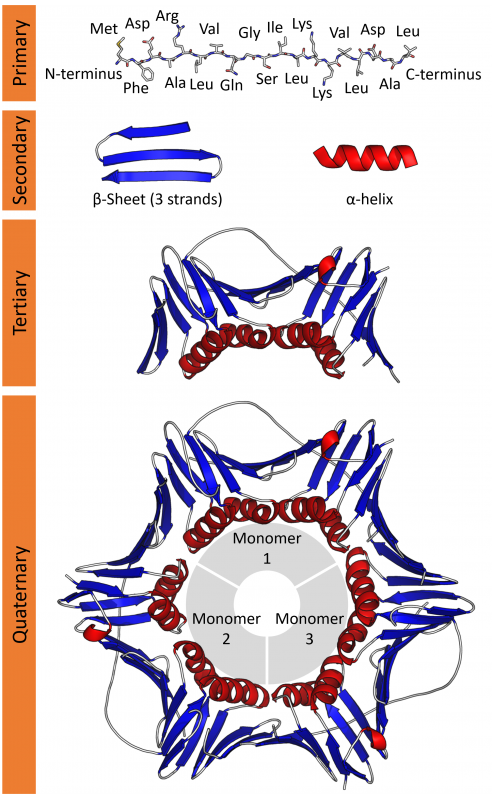 Organisms are made of molecules. This image illustrates the four "levels" of protein structure, from primary to quaternary. Toxins work in many different ways, but one of the simplest is to damage the structure of proteins, either at the tertiary level (resulting in the destruction of a single protein) or quaternary level (interfering with the interaction of sub-units that make up complex proteins). Image: Wikipedia.
Organisms are made of molecules. This image illustrates the four "levels" of protein structure, from primary to quaternary. Toxins work in many different ways, but one of the simplest is to damage the structure of proteins, either at the tertiary level (resulting in the destruction of a single protein) or quaternary level (interfering with the interaction of sub-units that make up complex proteins). Image: Wikipedia.
She has her father’s dental glands
All extant venomous reptiles are members of the Toxicofera (about which this tale is told!), and Toxicofera is comprised by Iguania, Anguimorpha, and Serpentes. The synapomorphy of Toxicofera is dental glands. “Synapo-what now?” A synapomorphy is a characteristic shared amongst all members of a particular clade (e.g. Toxicofera). It’s a very important concept in evolutionary biology because the inference we make when all members of a group of related species have a particular characteristic is that the ancestor of that group possessed that trait. Since the fossil record is patchy and not all traits fossilise well, we make inferences about what most recent common ancestors (MRCAs) probably looked like by looking at the shared traits of all the organisms that descend from them. Since all toxicoferan reptiles seem to have dental glands, we infer that the MRCA of Toxicofera had them too. Remember, dental glands differ from more typical oral glands in that they have ducts which deposit their secretions at the base of the teeth, instead of into the mouth more generally.

The dental glands of this bearded dragon (Pogona microlepidota) release their secretions at the base of its teeth. Image: Matt Summerville.
From iguanas to goannas
Iguanian lizards are, as the name suggests, iguanas and their relatives. In Australia, we have a huge diversity of “dragon lizards”, members of the family Agamidae, such as the bearded dragon, water dragon and frilled lizard. Agamid lizards are members of the Iguania, as are their relatives the chameleons (Chamaeleonidae). There are many other families in the Iguania, including several that are known as “iguanas” (another well-known group of iguanian lizards is the anoles). Iguanian lizards have dental glands on both the top and bottom jaws and the glands secrete both mucous and proteins. In this sense, they are like a typical “salivary gland”, in that they secrete substances that lubricate food and keep the mouth moist (mucous) and substances that aid in starting the digestion of food and contribute to oral hygiene (various proteins).
 Australia's iconic frilled lizard (Chlamydosaurus kingii) is a member of the family Agamidae, part of the Iguania. It is therefore (like the bearded dragon) a non-venomous member of the Toxicofera. Photo: Stewart Macdonald.
Australia's iconic frilled lizard (Chlamydosaurus kingii) is a member of the family Agamidae, part of the Iguania. It is therefore (like the bearded dragon) a non-venomous member of the Toxicofera. Photo: Stewart Macdonald.
Anguimorpha is a diverse clade of lizards – technically, it’s an infraorder, but that doesn’t really matter to us here. In Australia, the goannas (family Varanidae) fly the anguimorph flag. Australasia is the stronghold of the goannas, and over half the world’s species hail from our region. Another famous family of anguimorphs is Helodermatidae, which includes the gila monster (Heloderma suspectum) and Mexican beaded lizard (Heloderma horridum) – for a long time, these were considered the world’s only venomous lizards….or at least the only venomous lizards with legs (remember, snakes are lizards too!). There are six other families in the Anguimorpha, most of which only have one or a few species in them. The exception is Anguidae, which is quite diverse and includes the slow worms and their relatives; the galliwasps (I know, cool name right!?); and the alligator lizards. So far, most anguimorph lizards that have been examined only have dental glands on the lower jaw (mandibular dental glands). The one exception is the scheltopusik, a European legless lizard with the paradoxical scientific name Pseudopus apodus, which means “false foot no foot”.
It’s with the anguimorphs that dental glands start to get more interesting. In the dental glands of iguanian lizards, the serous-(protein) and mucous-secreting cells are mixed in together (although the concentration of mucous-secreting cells is higher towards the top of the gland and vice versa), making them similar to typical unspecialised oral glands. In the mandibular dental glands of some anguimorph lizards, however, these cells are segregated into specialised regions of the gland devoted solely to one type of secretion – lobules on top of the gland secrete only mucous, while lobules on the lower half of the gland secrete proteins. These regions are now so distinct as to almost be separate glands. Importantly, the protein-secreting cells dump their content into central “lumens” – receptacles in which the protein-rich solution can be stored, ready for release to the base of the teeth at very high concentrations. These specialised glands, possessed by anguimorph lizards from the families Varanidae (goannas or monitor lizards), Lanthanotidae (the Bornean earless monitor), and Helodermatidae (gila monster and beaded lizards), are starting to look a lot like “venom glands”.
 A heath monitor (Varanus rosenbergi). Monitor lizards (also known as "goannas") are members of the family Varanidae, part of the Anguimorpha. Whether or not monitor lizards are venomous remains controversial (and will be discussed in an upcoming AVRU blog post), but it's their impressive teeth that do most of the damage when they bite humans....humans silly and fast enough to get ahold of them! Photo: Matt Summerville.
A heath monitor (Varanus rosenbergi). Monitor lizards (also known as "goannas") are members of the family Varanidae, part of the Anguimorpha. Whether or not monitor lizards are venomous remains controversial (and will be discussed in an upcoming AVRU blog post), but it's their impressive teeth that do most of the damage when they bite humans....humans silly and fast enough to get ahold of them! Photo: Matt Summerville.
The snake situation
Snakes have a remarkable diversity of oral gland arrangements. For these limbless wonders that are notoriously able to subdue and swallow impressively large prey, oral glands have assumed central functional roles in feeding ecology. Of course, oral glands are always important for what goes into mouths (i.e. for feeding), but snakes seem to take this to the next level, which is perhaps why their oral glands are so diverse and (in some cases) so specialised. In fact, snakes exhibit such a plethora of transitional forms (such a litany of links!) amongst their oral gland arrangements, that a brief study of them is enough to refute the “missing links” argument of creationists and demonstrate the unfeasibility of Plato’s directive to “carve nature of the joints”. There’s something to be said about the influence of neo-Platonism on early Christian theology here, but perhaps a blog about venom systems is not the place to say it! Such a blog is also not the place for a detailed description of snake oral gland diversity, but the die-hard lovers of evolution should check out my paper “endless forms most beautiful” (a lovely turn of phrase borrowed from Darwin) to get the full run down (including many admissions of ignorance – we simply don’t know what some of these glands are really for!).
Unsurprisingly, the method by which a snake subdues its prey is strongly correlated with the development of its dental glands. At one end of the spectrum are the specialised constrictors – pythons, boas and numerous other related and unrelated species that have converged on this strategy. In those lineages that have been specialised constrictors for the longest period of time (pythons and boas), the protein-secreting component of the dental gland has receded into the corner of the mouth and decreased greatly in size (it’s now called a “rictal gland”, but this terminology becomes very confusing once you start digging). The major secretory structures near the teeth of these snakes are the labial glands, which aren’t really dental glands at all and almost exclusively secrete mucous. This makes sense, given that these snakes consume huge prey animals and need a lot of lubrication to get them down. In most lineages of snake, where protein-secreting dental glands are still in evidence, these are located on the upper jaw (they are maxillary dental glands). There are a couple of fascinating exceptions – snakes which have developed toxin-secreting dental glands on the mandible – but almost all snakes have the opposite arrangement to that of the anguimorph lizards described above. Why the anguimorphs went one way and the snakes the other, no one knows – in evolution, whatever can happen will happen, and it may be that this divergence was entirely random.
A continuum of categories
As you may have guessed at this point, the maxillary dental glands of snakes include the bona fide “venom glands” of front-fanged snakes like vipers and elapids. These snakes are at the opposite end of the spectrum from the constrictors, in that they primarily use venom for subduing prey (although many aren’t shy about throwing a coil or two around a prospective meal). The diversity of venom glands and quasi venom glands in snakes is so rich and fascinating that we’re going to have to wait until next week to dig into it. We’ll see that this diversity, and an innate desire to be proper Platonists by grouping these forms into discrete categories, has led to terminological confusion and a lot of debate about what exactly a “venom gland” is, anyway. Cutting to the chase, I’d like to point out ahead of time that a venom gland is a gland that secretes venom (!). Venom is a functional trait (remember?) and thus venom glands constitute a function category…..oh dear, it may be harder to avoid carving nature into categories than we hoped! It’s true, we do need to classify things, and no classificatory scheme is perfect – it’s a map, not the territory. This is as true of musical genres as it is of biological nomenclature (systems of naming). Classification is useful and necessary, and despite the fact that no scheme is perfect, some are better than others. It may even turn out that there is no “one size fits all” scheme – perhaps it depends on the question we’re trying to ask or the concept we’re trying to communicate. Luckily, we’re not in it alone; we work out which words are fit for purpose in a constant negotiation with other people in the “space of reasons”. This is what scientists and philosophers are doing when they debate the nature of reality and the meanings of the words we use to describe it – they’re negotiating. We’re all part of this process, because language evolves as a consequence of patterns of usage and culture evolves in part as a consequence of the way in which patterns of language usage inform the content of worldviews.

Although this eastern brown snake (Pseudonaja textilis) is of course highly venomous, it has also wrapped several coils around the unfortunate blue-tongued skink (Tiliqua scincoides) it is trying to subdue. Whether this behaviour is truly "constriction", or rather an attempt to keep the lizard still enough for the snake to work one of its small stiff fangs in-between the lizard's bony scales, remains poorly understood. Photo: Matt Clancy.
Tune in next week for more evolutionary thinking and a closer look at the venom glands of snakes, as well as the spectrum of venom systems among toxicoferan reptiles, from the “incipient” (at an early stage of development) to the advanced.
- Timothy