From toxins to t-cells, in evolution existence precedes essence
“Existence precedes essence”, the credo of Existentialist philosophy, is also a central tenet of evolutionary thinking.
Existentialism
The French philosopher Jean-Paul Sartre coined the phrase “existence precedes essence” to describe the core proposition of a strain of philosophy (Existentialism) that preceded his own existence by centuries, if not millennia. What does this deceptively simple assertion actually mean? No more (or less) than that humans do not come into being with a ready-made purpose or meaning (an “essence”) but have to create their own reason for being through their actions and their understanding of those actions. We tell our own stories; justify our own existence. For Sartre, this was a criticism of religious views in which the meaning of life is imposed on us from outside – by God or gods. He rejected this notion, insisting that we must create our own unique meaning of life, formulated within ourselves and then enacted in the world. This meant that humans were radically free, but with this freedom came a great deal of responsibility – time to grow up and make our own beds. Whilst this view is very popular today it’s also not beyond reproach, and it would be quite possible (and fun) to spend several paragraphs criticising it. On the other hand, this is not a philosophy blog – it’s a venom biology blog – so the onus is now on me to justify bringing it up at all. What exactly does “existence precedes essence” have to do with biology?!
 Jean-Paul Sartre (1905-1980) was one of the 20th Century's most popular and influential writers and philosophers. He is pictured here with his life companion and fellow philosophical luminary, author of The Second Sex, Simone de Beauvoir (1908-1986). Sartre coined the term "existence precedes essence" in his essay "Existentialism is a Humanism" which was aimed at dispelling the notion that Existentialism was a gloomy or pessimistic philosophy. Photo: Getty Images.
Jean-Paul Sartre (1905-1980) was one of the 20th Century's most popular and influential writers and philosophers. He is pictured here with his life companion and fellow philosophical luminary, author of The Second Sex, Simone de Beauvoir (1908-1986). Sartre coined the term "existence precedes essence" in his essay "Existentialism is a Humanism" which was aimed at dispelling the notion that Existentialism was a gloomy or pessimistic philosophy. Photo: Getty Images.
Exaptation
It turns out that this notion that things (all kinds of things) first emerge for no particular reason, and then “discover” their reason for being retroactively, is a core part of the view of the world provided by the lens of contemporary evolutionary biology. I’ve discussed the phenomenon of “exaptation” in a few blog posts already (e.g. here) – it’s what we call the process in which a property (something that merely exists) becomes a function (something that has a reason for existing – a purpose). The term was coined in 1982 by the palaeontologists Stephen Jay Gould and Elisabeth Vrba. Gould was an extremely famous and influential evolutionary biologist and in science fame and influence are almost invariably accompanied by controversy. Gould had numerous intellectual feuds with other influential biologists, such as Richard Dawkins and Simon Conway Morris, and most ideas associated with him have been extensively criticised. Many of them have become canonical, however, including exaptation. It’s not that Gould and Vrba were the first to realise that properties become functions, but they were the first to give the process a name. The philosopher Daniel Dennett once observed that exaptation was a “trivial” (obvious, barely worth drawing attention to) process, since all adaptations begin as exaptations. Although I greatly admire Dennett, he misses the point here – it’s precisely because exaptation is trivial and ubiquitous that people completely overlook it, and thus why it required a catchy name. We mustn’t forget that evolution itself is equally trivial – it’s just future states of systems being descended from past states – and that Thomas Huxley, speaking of Darwin’s monumental “discovery” of natural selection, remarked that it was “extremely stupid not to have thought of that!”. The best ideas are often those that seem most obvious as soon as someone else has identified them.

Stephen Jay Gould (1941-2002) was a palaeontologist and one of the most influential evolutionary biologists of the 20th Century. Whilst many of his ideas remain controversial within the field, his impact upon it is unquestionable. Some of his most famous contributions include the idea of "punctuated equilibrium" - the assertion that evolution typically proceeds slowly but occasionally leaps forward; his cautions against "adaptationism" - the notion that all of an organism's features are adapted and functional (rather than some being simply "properties"); and exaptation. Image: Newsweek.
Order from randomness
It's also the case that the simplest and most ubiquitous of processes can have the most counter-intuitive of consequences. One of the most difficult things to understand about evolution is the way in which apparently “random” change ends up creating all the wondrous order and complexity of the natural world. This has long been one of the most powerful arguments in support of creationism – “surely you don’t believe that all this beauty, all this structure, could be the result of a random process? In order for complex structure to emerge, it takes an even more complex process to design it!”. Well, sure – I don’t believe it’s the result of a random process, I believe it’s the result of a process that turns more or less random change into absolutely non-random structure. The change isn’t “absolutely” random, either, it’s just undirected – change simply happens and changes become purposeful (or not) after they emerge. Existence precedes essence – that’s exaptation. I also agree with the second point – evolution is a highly complex process, more complex than any of its products, but that complexity is the result of iterated (repeated) feedback cycles, it’s not all there “in the beginning”. Complexity accumulates as the result of the repetition of an extraordinarily simple process of change and constraint – change occurs (more or less randomly) and that change is constrained by what already exists (the past constrains the future). As complex structure forms, the potential emerges for more complexity to be built atop what is already present. It’s important to note here that “evolution” is the whole process – the random change and the constraint. Exaptation is part of that process – the conversion of random change into purposeful product via the mechanism of selection (there’s more to the concept than this, but I’ve discussed that in other AVRU blogs). Selection is one form of constraint on change – it’s the kind that Darwin identified and it’s extremely important for the evolution of biological organisms.

William Paley (1743-1805) was an English clergyman and philosopher. He is most famous for his use of the "watchmaker analogy" in the book Natural Theology, or Evidences of the Existence and Attributes of the Deity collected from the Appearances of Nature. The gist of the analogy is that if one were to stumble across a watch - an intricately designed artefact - just lying about, one would automatically assume that something so complex must have been created by an intelligent designer. Following the same intuition, Paley says that we should assume the same conclusion when we observe the complexity of nature. The book was published in 1802, 57 years before Darwin published On the Origin of Species - arguments between proponents of an intelligent designer and those of an impersonal evolutionary process are ancient, and did not begin (or end) in the Victorian Era. Richard Dawkins referenced Paley's argument in his book The Blind Watchmaker. Painting by George Romney.
Innate and adaptive immunity
OK, that’s more than enough theory for now, how about some examples? It’s probably safe to say that immunity has been on a lot of people’s minds lately. We’d all like to be immune to the coronavirus responsible for Covid-19. But how does immunity work? There are two basic kinds of immunity – innate and adaptive (acquired). Innate immunity is something organisms are born with – it’s conferred by their genes. As AVRU blog readers know, organisms are fantastically elaborate chemistry sets – we produce a huge variety of functional molecules, like proteins, that are responsible for all sorts of activities within our bodies. A subset of these molecules was designed (by natural selection!) to protect us from invaders. For example, we produce enzymes that are great at killing bacteria. Since we produce these innately, they are ready to go at any time and are thus our first line of defence against invaders. They’re quite relevant to toxin evolution, because some enzymes that have been recruited for use as toxins are ancestrally antibacterial. You may recognise this as an example of exaptation – the molecules had one function, which made them good at killing cells (e.g. bacteria) and this function made them potentially useful for injecting into other organisms in order to subdue them. Thus, phospholipases, which are ancestrally antibacterial and part of our innate immune response, have been recruited as toxins by a number of venomous lineages (e.g. snakes and bees).

Phospholipases, which ancestrally functioned as components of the innate immune response, are important toxins in the venom of the Mojave rattlesnake (Crotalus scutulatus) and other vipers. The Mojave rattlesnake is considered one of the most dangerous of all rattlesnakes, due to the presence of a neurotoxin in its venom, formed from two phospholipase enzymes stuck together to form a complex toxin. Photo: BLM Nevada, Wiki Commons.
Unfortunately, we don’t have innate immunity against coronaviruses. So how does adaptive immunity work? Not only is the process of acquiring immunity to unique pathogens extremely important to our survival, it’s exceptionally cool and is a beautiful microcosm of the evolutionary process in which undirected change is converted into functional products. B-cells are a type of white blood cell that secretes antibodies. When a B-cell comes in contact with an “antigen” (something that stimulates an immune response) it begins to multiply by dividing itself over and over again, and this is where the fun begins. During this process of proliferation, “somatic hypermutation” occurs in the aptly named “hypervariable” regions of the antibodies that the B-cell produces. These are the regions of the antibodies that detect and bind to antigens. As you know, “hyper” means “a lot”, so what this means is that a great deal of mutation is occurring in these regions – the mutation rate may be 1 million times higher than normal – resulting in, well, a lot of variation! Now, you’ve probably guessed the punchline here – all this variation is undirected. It’s essentially random (although of course where it occurs is not random at all). Most of this variation is completely meaningless and doesn’t result in an increased affinity (ability to bind) for the antigen. It’s a bit like scrolling randomly through channels on the radio (if anyone remembers radio) until you hit upon something you like. By chance, some small subset of all that change in the binding regions of the antibodies increases their affinity for the antigen, and when that happens, that undirected change has found its calling – it now has a purpose. Note that the same variations may have come up many times before and remained meaningless – meaning (function) only emerges when the right change occurs at the right time. Meaning is dependent on the relationship between the change in the variable region and the structure of the antibody. When a match occurs by chance, selection occurs – the B-cells with the right antibodies start to proliferate preferentially and we’re on our way to an adaptive immune response. This is exactly what happens in evolution in general – undirected mutations largely have meaningless (typically neutral) or occasionally deleterious results. On occasion, the right mutation occurs at the right time and positive selection occurs – a novel function is born. Most change has results that are “epiphenomenal” (another way of saying it “just is”), but when a change encounters the right context, it is exapted and becomes functional.
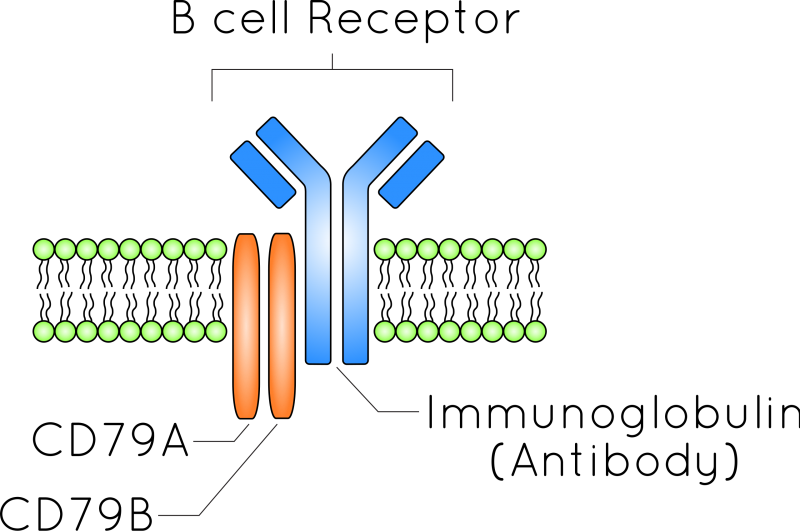
The hypervariable regions of the B cell receptor - which is a transmembrane antibody that functions as an antigen receptor - undergo somatic hypermutation. The mutation rate of these regions may be as much as 1 million times greater than the background rate, and they continue changing until they hit upon the solution to a problem - effective binding of a pathogenic invader. Image: Wikipedia.
Memory T-cells and Covid-19
OK, now I have to justify the alliteration in the title – it mentions T-cells, not B-cells. Well, T-cells are another kind of white blood cell, and one of the things they do is kill infected cells directly. For a long time it was thought that somatic hypermutation only occurred in B-cells, but more recently evidence has emerged to suggest that it occurs in T-cells too (at least in camels and sharks). This is obviously cool from an evolutionary perspective, but T-cells have also been in the news recently for slightly different reasons. T-cells multiply in response to the detection of a pathogen, but after the infection is dealt with most of the T-cells specific to that pathogen die. A small subset – the “memory T-cells” – remains, however, as part of a persistent acquired immune response to a particular pathogen. Next time you encounter that “bug”, you’re able to respond to it more rapidly. Remember SARS (severe acute respiratory syndrome) from back in 2003? It was caused by a coronavirus. Turns out that the memory T-cells of patients that recovered from SARS infection also recognise the coronavirus that causes Covid-19 (check out the study here). In fact, it looks like a surprisingly high percentage of the population, with no exposure to the SARS or Covid-19 viruses, might already have memory T-cells that recognise these potential invaders (see additional studies here and here). This could be part of the reason why some people have relatively mild reactions to the virus. From the perspective of this article, it’s another incredible example of exaptation. These memory T-cells that recognise the Covid-19 virus did not evolve for that purpose. In fact, they evolved (or were developed, if you prefer) before the virus itself even existed! This means that the particular variants of the surface proteins of the T-cells were meaningless (epiphenomenal) until they met the surface proteins of the virus. Many years after they came into existence, these T-cells discovered their essence.
Evolution is just so cool
Since this kind of thing is ubiquitous in evolution, it probably goes without saying that similar things happen with toxins. Sometimes the possibility of a molecule becoming a toxin doesn’t exist until it is accidentally injected into another organism. For example, antibacterial molecules in lizard saliva – they weren’t designed to weaken prey animals, they just got injected into those prey animals as part of a typical predatory interaction in which the lizard was trying to subdue its meal by biting it. Sometimes those molecules have properties that have no function whatsoever, but those properties suddenly become functional when they are introduced to a new context – like the insides of another animal. That’s existence preceding essence. Exaptation.
In a future article, we’re going to explore the ways in which toxins become toxins – “how the toxin got its toxicity” – in more depth. There’s also another exciting (I mean, really really exceptionally fantastically cool) thing to talk about in relation to adaptive immunity. Remember I said that the process of somatic hypermutation was like a microcosm of the evolutionary process? The thing is, in the case of adaptive immunity, the mechanics of evolution have themselves been selected for – it’s a functional exploitation of the process of evolution itself! Not sure why that’s mind-bogglingly cool? Tune in to future AVRU blog posts in which I’ll try to convince you that your jaw should be on the floor…..